DNA Replication (DNA 複製)
| 階段 | 比較項目 | 原核生物 (E. coli) | 真核生物 |
|---|---|---|---|
| 起始 (Initiation) |
辨認 起始位點 |
DnaA 結合至 oriC 序列 (245 bp, AT-rich) |
ORC (Origin Recognition Complex) 結合至 ARS |
| 解旋 (Helicase) |
DnaB (DnaC 協助) |
MCM complex (2-7) (CDC6 / CDT1 協助) |
|
| 引子合成 (Primase) |
DnaG 合成 RNA primer |
DNA Pol α | |
| 拓撲結構 解鎖 |
Gylase (Topo II) 減少 (+),增加 (-) 超螺旋 |
Topo I、Topo II | |
| 防止 再複製 |
Dam methylase: OriC 的 5'-G==ATC (MMR 後) (Adenine-N6== methylation) |
Geminin 封鎖 CDT1 防止 Pre-RC 再次組裝 |
|
| 延長 (Elongation) |
主要 合成酶 |
DNA Pol III | DNA Pol ε (L==e==ading) DNA Pol δ (Lagging) |
| 單鏈保護 | SSB (ssDNA binding protien) |
RPA (Replication protein A) |
|
| 滑動鉗 | β-clamp (γ -complex 裝載) |
PCNA (RFC 裝載) |
|
| 連接 岡崎片段 |
DNA Ligase (需 NAD+): 具 Lys,需 AMP 共價催化 (Adenylylation of DNA Ligase) |
DNA Ligase (需 ATP): 催化模式同原核 |
|
| 引子移除 | DNA pol I (移除 + 填補) | 移除:RNase H + FEN1 填補:DNA Pol δ |
|
| 終止 (Termination) |
終止位點 | Tus-Ter complex: ==Tus 蛋白==結合 Ter sequences 在 Ter site 阻擋複製叉 |
無特定終止序列 複製叉相遇即融合 |
| 拓撲結構 解鎖 |
Topo IV 分離複製完成的兩組 DNA (catanane) |
Topo II 協助線性染色體分離 |
|
| 末端問題 | 無 (環狀 DNA 無末端) | Telomere 縮短 需 Telomerase 修復 |
-
Pre-RC 組裝:
- 只在 G1 phase,ORC, CDC6, CDT1, 協助兩個 MCM2-7 組裝
- 組裝過程為 ATP-dependent:
- ORC, CDC6, MCM 屬於 AAA+ATPase family
- ORC, CDC6 具有 molecular chaperone 功能 → 協助 MCM 正確組裝
- MCM 組裝正確後,ATP 才可順利水解,使裝載完成,啟動後續流程
- 共同組成 Pre-Pc double Hexamer
-
Pre-RC 活化:
- 在 S 期被 S-CDK 等磷酸化
- MCM2-7 活化,解旋 DNA,開啟 elongation
-
阻止 Pre-RC 再組裝:
- 避免 DNA 重複複製
- S-CDK:磷酸化 CDC6 / CDT1 使其降解、離開細胞核
- Geminin:
- 封鎖 CDT1 (MCM 穩定嚮導),使其無法與 MCM 結合
- Geminin 只在 G1 不活化 → Pre-RC 只能在 G1 phase 組裝
-
DNA 複製四大特性:
-
Semiconservative (半保留):
- 分離的兩股各自作為新股模板,合成新股
- 利用==無放射性的 15N== 驗證
- 複製前後懸浮位置不同:15N-15N 最重,15N-14N 次之,14N-14N 最輕
-
Bidirectionally (雙向複製):
- 從複製起點 Origin (Ori) 解旋產生複製泡
- Origin (Ori) 位於複製泡
- Ori 為 AT-rich 序列,氫鍵較少,容易解旋
- 兩端形成動態分叉口 (Replication fork) 分頭複製
- 從複製起點 Origin (Ori) 解旋產生複製泡
-
5' → 3' Direction (方向性):
- 新股合成的方向固定由 5' → 3'
- 上一個 dNTP 的 3'-OH 攻擊下一個 dNTP 的 5'-α-Phosphate
- 形成 Phosphodiester bond
- 剩下的 Biphosphoate (β, γ) 將被 Pyrophosphatase 分解成 2 Pi
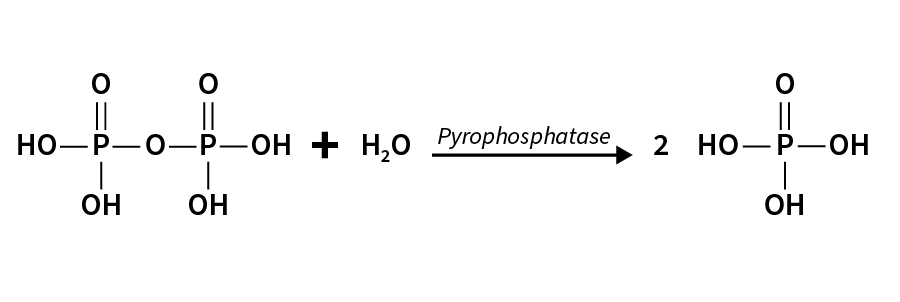
- Pyrophosphatase 可協助推動 DNA / RNA 合成 (使 rxn 向右)
-
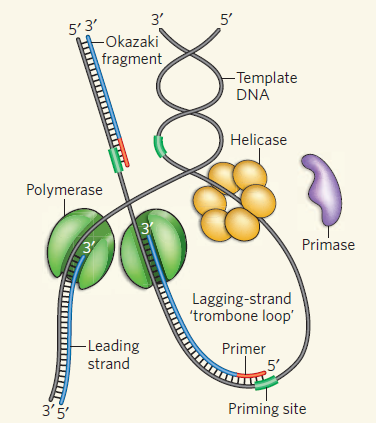
Semidiscontiuous (半不連續性):
- 為遵守方向性,一股連續合成,一股分段合成
- Leading strand:
- 連續合成,又稱 contiuous strand
- 合成==較早開始,但與延遲股合成速度相同==
- Lagging strand:
- 分段合成,又稱 discontiuous strand
- 合成==較晚開始,但與領先股合成速度相同==
- Leading strand:
- Okazaki fragments:
- Lagging strand 的合成片稱為岡崎片段
- 所有生物皆有 (病毒無)
- 為遵守方向性,一股連續合成,一股分段合成
-
- Protein-coding gene:
- Exon:佔比 1.5 %
- Non-protein-coding gene:
- Intron:佔比 25 %
- Transposon:佔比 45 %
- Class I: Retrotransposons (42%)
類似困在細胞內的病毒,目的是增加自己的 gene copy- LINEs
- SINEs:Alu elements 屬於此類
- LTR retrotransposons
- Class II:DNA transposons (3%)
- Class I: Retrotransposons (42%)
| 類別 | 移動方式 | 關鍵酵素 | 結構特徵 / 備註 |
| :------------------------: | :------------------: | :----------------------------------: | :-----------------------------------------------------: |
| LTR | Copy & Paste | RTase + Integrase | 具 Long Terminal Repeats |
| LINEs | Copy & Paste | RTase + Endonuclease | mRNA-like,由 RNA Pol II 轉錄
利用 TPRT 機制 |
| SINEs | Copy & Paste | 借用 LINEs 的酵素 | tRNA-like,由 RNA Pol III 轉錄 |
| DNA
Transposon | Cut & Paste | Transposase | 兩端具 Inverted Repeats (IR)
(transposase 剪切點) |
-
原核生物 DNA 複製 (E.coli):
| 階段 | 原核生物 (E. coli) | 功能 |
|---|---|---|
| 起始 (Initiation) |
DnaA | 辨認起始位點:結合 ori 序列 (245 bp, AT-rich) |
| DnaB | Helicase:解旋 DNA (由 DnaC 協助) | |
| DnaG | Primase:合成 RNA primer | |
| Gylase (Topo II) | 拓撲結構解鎖:增加 (-) supercoil,減少 (+) | |
| Dam methylase |
防止再複製、區分新舊股: Ori 的 5'-G==ATC 甲基化 (Adenine-N6== methylation) |
|
| 延長 (Elongation) |
DNA Pol III | 主要合成酶 (多次單元) β-clamp 在 DNA 上滑動 (clamp 由 γ -complex 裝載) |
| SSB (ssDNA binding protien) |
單鏈保護:保護 ssDNA 結構 | |
| DNA Ligase | 連接岡崎片段: 具 Lys,需 AMP 共價催化 (需 NAD+) (Adenylylation of DNA Ligase) |
|
| DNA pol I | 引子移除 | |
| 終止 (Termination) |
Tus-Ter complex | 終止複製:==Tus 蛋白==結合 Ter sequences 在 Ter site 阻擋複製叉 |
| Topo IV | 分離複製完成的兩組環狀 DNA (catanane 結構) |
| 特徵 | DNA Pol I | DNA Pol II | DNA Pol III |
|---|---|---|---|
| 5'→3' Polymerase (DNA 聚合) |
有 | 有 | 有 |
| 3'→5' Exonuclease (Proofreading) |
有 | 有 | 有 |
| 5'→3' Exonuclease (Primer 切除) |
有 | 無 | 無 |
| 主要角色 | 移除 primer 置換為 DNA |
DNA 修復 重啟複製 |
DNA合成主力 |
| Processivity (一次結合之合成數量) |
低 (切完 primer 就掉下來) |
中 | 極高 (一次跑幾萬個 bp) |
| 分子結構 | 單一肽鏈 三級結構 |
單一肽鏈 三級結構 |
多次單元,四級結構 Holoenzyme |
| 功能區塊 | Subunits | 主要功能 (Function) |
| :------------------------------------: | :-----------------------------------: | :------------------------------------------------------------: |
| Core Enzyme |
| |
| |
| β-clamp
(Sliding Clamp) |
β-clamp 把酶鎖在 DNA 上,防止脫落 |
| γ-complex |
| Dimerization |
- DNA Pol I 經蛋白酶切割可分成大小兩片段,Nick translation 需整合兩片段的活性
- Nick translation 包含 Primer 移除 + DNA 加入,單一片段不具有 Nick translation 能力
- | 片段名稱 | 包含端點 | 包含活性 | Domain 功能 |
| --------------------------------------------------- | ------------------ | -------------------------- | --------------------------- |
| Small Fragment | N-terminal | - 5'→3' Exo | Primer 移除 |
| Large Fragment
(Klenow Fragment) | C-terminal | - 5'→3' Pol
- 3'→5' Exo | 聚合:加入核苷酸
校正:Proofreading | - 每一個 Okazaki fragment 的 5' 端皆有 RNA primer,此處的 DNA 由 Pol I 合成
- Pol I 邊切除 primer,邊加入 DNA → Nick 移動
- 5'-|ddddddd| |rrrrrrrrr|dddddddddddddddd|-3
- 5'-|dddddddddd| |rrrr|dddddddddddddddd|-3'
- 5'-|ddddddddddddd| |dddddddddddddddd|-3'
- 真核、原核皆使用此模型
- DNA 的 Leading / Lagging strand 各自有 Core Enzyme 催化
- 兩 Core enzymes 透過
subunit 連結成 dimer,同時催化 DNA 合成 - 藉由 Lagging strand looping (形成迴圈),使其與 leading strand 能「同向」合成 DNA
- Leading strand:連續合成
- Helicase 在前方解旋,Core enzyme 順順的向前合成,不需反覆裝載
- Lagging strand:重複換軌 (clamp)
- 合成一段岡崎片段後,Core enzyme 撞到前一段合成的片段
- 構型改變導致其與原本的
-clamp 的親和力下降 - Core enzyme 自動鬆脫,快速回彈至下一個預裝好的 clamp 處,繼續下一段複製